
Innenohr - Inner ear
| Innenohr | |
|---|---|
 | |
| Einzelheiten | |
| Arterie | Labyrintharterie |
| Kennungen | |
| Latein | auris interna |
| Gittergewebe | D007758 |
| TA98 | A15.3.03.001 |
| TA2 | 6935 |
| FMA | 60909 |
| Anatomische Terminologie | |
 |
| Dieser Artikel gehört zu einer Reihe, die die Anatomie der |
| Menschliches Ohr |
|---|
|

Das Innenohr ( Innenohr , auris interna ) ist der innerste Teil des Vertebraten ear . Bei Wirbeltieren ist das Innenohr hauptsächlich für die Geräuscherkennung und das Gleichgewicht verantwortlich. Bei Säugetieren besteht es aus dem knöchernen Labyrinth , einer hohlen Höhle im Schläfenbein des Schädels mit einem Durchgangssystem, das zwei Hauptfunktionsteile umfasst:
- Die Cochlea , die dem Hören gewidmet ist; Umwandlung von Schalldruckmustern vom Außenohr in elektrochemische Impulse, die über den Hörnerv an das Gehirn weitergegeben werden .
- Das Vestibularsystem , das dem Gleichgewicht gewidmet ist
Das Innenohr ist bei allen Wirbeltieren zu finden und weist erhebliche Unterschiede in Form und Funktion auf. Das Innenohr wird bei allen Wirbeltieren vom achten Hirnnerv innerviert .
Struktur

Das Labyrinth kann nach Schichten oder Regionen unterteilt werden.
Knochen- und Membranlabyrinthe
Das knöcherne Labyrinth oder knöcherne Labyrinth ist das Netzwerk von Passagen mit knöchernen Wänden, die mit Periost ausgekleidet sind . Die drei Hauptteile des knöchernen Labyrinths sind das Vestibül des Ohrs , die halbkreisförmigen Kanäle und die Cochlea . Das membranöse Labyrinth verläuft innerhalb des knöchernen Labyrinths und erzeugt drei parallele, mit Flüssigkeit gefüllte Räume. Die beiden äußeren sind mit Perilymphe und die innere mit Endolymphe gefüllt.
Vestibular- und Cochlea-Systeme
Im Mittelohr wird die Energie der Druckwellen von den drei Gehörknöchelchen in mechanische Schwingungen umgewandelt. Druckwellen bewegen das Trommelfell, das wiederum den Malleus, den ersten Knochen des Mittelohrs, bewegt. Der Malleus artikuliert zum Incus, der mit den Stapes verbunden ist. Die Fußplatte der Stapes verbindet sich mit dem ovalen Fenster, dem Anfang des Innenohrs. Wenn der Stapes auf das ovale Fenster drückt, bewegt sich die Perilymphe, die Flüssigkeit des Innenohrs. Das Mittelohr dient somit dazu, die Energie von Schalldruckwellen in eine Kraft auf die Perilymphe des Innenohrs umzuwandeln. Das ovale Fenster hat nur ungefähr 1/18 der Fläche des Trommelfells und erzeugt somit einen höheren Druck . Die Cochlea verbreitet diese mechanischen Signale als Wellen in der Flüssigkeit und in den Membranen und wandelt sie dann in Nervenimpulse um, die an das Gehirn übertragen werden.
Das Vestibularsystem ist der Bereich des Innenohrs, in dem die halbkreisförmigen Kanäle nahe der Cochlea zusammenlaufen. Das Vestibularsystem arbeitet mit dem visuellen System zusammen, um Objekte im Blick zu behalten, wenn der Kopf bewegt wird. Gelenk- und Muskelrezeptoren sind ebenfalls wichtig für die Aufrechterhaltung des Gleichgewichts. Das Gehirn empfängt, interpretiert und verarbeitet die Informationen aus all diesen Systemen, um das Gefühl des Gleichgewichts zu erzeugen.
Das Vestibularsystem des Innenohrs ist für die Gleichgewichts- und Bewegungsempfindungen verantwortlich. Es verwendet die gleichen Arten von Flüssigkeiten und Detektionszellen ( Haarzellen ) wie die Cochlea und sendet Informationen über die Haltung, Rotation und lineare Bewegung des Kopfes an das Gehirn. Die Art der Bewegung oder Haltung, die von einer Haarzelle erfasst wird, hängt von den zugehörigen mechanischen Strukturen ab, wie z. B. dem gekrümmten Rohr eines halbkreisförmigen Kanals oder den Calciumcarbonatkristallen ( Otolith ) des Sacculus und des Utrikels .
Entwicklung
Das menschliche Innenohr entwickelt sich während der vierten Woche der Embryonalentwicklung aus dem auditorischen Placode , einer Verdickung des Ektoderms, die die bipolaren Neuronen der Ganglien Cochlea und Vestibularis hervorruft . Wenn der auditive Placode in Richtung des embryonalen Mesoderms invaginiert , bildet er das auditive Vesikel oder die Otozyste .
Aus dem auditorischen Vesikel entstehen die utriculären und sacculären Komponenten des membranösen Labyrinths . Sie enthalten die sensorischen Haarzellen und Otolithen der Utrikelmakula bzw. des Sacculus , die auf die lineare Beschleunigung und die Schwerkraft reagieren . Die utriculäre Teilung des auditorischen Vesikels reagiert auch auf die Winkelbeschleunigung sowie auf den endolymphatischen Sack und den Gang , die den Sacculus und den Utrikel verbinden.
Ab der fünften Entwicklungswoche entsteht aus dem auditorischen Vesikel auch der Cochlea-Gang , der das Spiralorgan von Corti und die Endolymphe enthält , die sich im membranösen Labyrinth ansammelt. Die vestibuläre Wand trennt den Cochlea-Gang von der perilymphatischen Scala vestibuli , einer Höhle innerhalb der Cochlea. Die Basilarmembran trennt den Cochlea-Gang von der Scala Tympani , einem Hohlraum innerhalb des Cochlea-Labyrinths. Die Seitenwand des Cochlea-Ganges wird vom Spiralband und der Striaascularis gebildet , die die Endolymphe produziert . Die Haarzellen entwickeln sich aus den lateralen und medialen Graten des Cochlea-Ganges, die zusammen mit der Tektorialmembran das Corti-Organ bilden.
Mikroanatomie


Der Rosenthal-Kanal oder der Spiralkanal der Cochlea ist ein Abschnitt des knöchernen Labyrinths des Innenohrs, der ungefähr 30 mm lang ist und 2¾ Umdrehungen um den Modiolus macht , die Mittelachse der Cochlea, die das Spiralganglion enthält .
Zu den spezialisierten Innenohrzellen gehören: Haarzellen, Säulenzellen, Boettcher-Zellen, Claudius-Zellen, Spiralganglion-Neuronen und Deiters-Zellen (Phalangealzellen).
Die Haarzellen sind die primären auditorischen Rezeptorzellen und sie sind auch als auditive Sinneszellen, akustische Haarzellen, auditive Zellen oder Zellen von Corti bekannt. Das Corti-Organ ist mit einer einzelnen Reihe innerer Haarzellen und drei Reihen äußerer Haarzellen ausgekleidet. Die Haarzellen haben ein Haarbündel an der apikalen Oberfläche der Zelle. Das Haarbündel besteht aus einer Reihe von Stereozilien auf Aktinbasis. Jedes Stereocilium fügt sich als Wurzel in ein dichtes filamentöses Aktin-Netz ein, das als Kutikularplatte bekannt ist. Eine Störung dieser Bündel führt zu Hörstörungen und Gleichgewichtsstörungen.
Innere und äußere Säulenzellen im Corti-Organ unterstützen Haarzellen. Äußere Säulenzellen sind einzigartig, da es sich um freistehende Zellen handelt, die nur benachbarte Zellen an den Basen und Spitzen berühren. Beide Arten von Säulenzellen weisen Tausende von vernetzten Mikrotubuli und Aktinfilamenten in paralleler Ausrichtung auf. Sie sorgen für eine mechanische Kopplung zwischen der Basalmembran und den Mechanorezeptoren an den Haarzellen.
Boettcher-Zellen befinden sich im Corti-Organ, wo sie nur in der unteren Windung der Cochlea vorhanden sind. Sie liegen auf der Basilarmembran unter Claudius 'Zellen und sind in Reihen angeordnet, deren Anzahl zwischen den Arten variiert. Die Zellen interdigitalisieren miteinander und projizieren Mikrovilli in den Interzellularraum. Sie unterstützen Zellen für die auditorischen Haarzellen im Corti-Organ. Sie sind nach dem deutschen Pathologen Arthur Böttcher (1831-1889) benannt.
Claudius 'Zellen befinden sich im Corti-Organ, das sich über Reihen von Boettcher-Zellen befindet. Wie Boettchers Zellen gelten sie als Stützzellen für die auditorischen Haarzellen im Corti-Organ. Sie enthalten eine Vielzahl von Aquaporin- Wasserkanälen und scheinen am Ionentransport beteiligt zu sein. Sie spielen auch eine Rolle bei der Abdichtung endolymphatischer Räume. Sie sind nach dem deutschen Anatom Friedrich Matthias Claudius (1822-1869) benannt.
Deiters-Zellen (Phalangealzellen) sind eine Art von Neurogliazellen , die im Corti-Organ vorkommen und in einer Reihe innerer Phalangealzellen und drei Reihen äußerer Phalangealzellen organisiert sind. Sie sind die Stützzellen des Haarzellenbereichs innerhalb der Cochlea. Sie sind nach dem deutschen Pathologen Otto Deiters (1834-1863) benannt, der sie beschrieb.
Hensens Zellen sind hochsäulige Zellen, die direkt an die dritte Reihe von Deiters-Zellen angrenzen.
Hensens Streifen ist der Abschnitt der Tektorialmembran über der inneren Haarzelle.
Nuels Räume beziehen sich auf die mit Flüssigkeit gefüllten Räume zwischen den äußeren Säulenzellen und benachbarten Haarzellen sowie auf die Räume zwischen den äußeren Haarzellen.
Die Hardesty-Membran ist die Schicht der Tectoria, die der retikulären Lamina am nächsten liegt und über der äußeren Haarzellregion liegt.
Die Reissner-Membran besteht aus zwei Zellschichten und trennt das Scala-Medium vom Scala-Vestibuli.
Huschkes Zähne sind die zahnförmigen Grate am Spiral-Limbus, die mit den Tectoria in Kontakt stehen und durch Interdentalzellen getrennt sind.
Blutversorgung
Das knöcherne Labyrinth erhält seine Blutversorgung aus drei Arterien: 1. Vorderer Trommelfellast (aus der Arteria maxillaris). 2- Petrosalast (von der mittleren Meningealarterie). 3- Stylomastoider Ast (von der A. auricularis posterior). Das membranöse Labyrinth wird von der Labyrintharterie versorgt . Die venöse Drainage des Innenohrs erfolgt durch die labyrinthische Vene, die in den Sigmoid-Sinus oder den unteren Petrosal-Sinus mündet .
Funktion
Neuronen im Ohr reagieren auf einfache Töne, und das Gehirn dient dazu, andere zunehmend komplexe Geräusche zu verarbeiten. Ein durchschnittlicher Erwachsener kann normalerweise Geräusche zwischen 20 und 20.000 Hz erkennen. Die Fähigkeit, höhere Tonhöhen zu erkennen, nimmt bei älteren Menschen ab.
Das menschliche Ohr hat sich mit zwei grundlegenden Werkzeugen entwickelt, um Schallwellen zu codieren. Jedes ist bei der Erkennung von hoch- und niederfrequenten Geräuschen getrennt. Georg von Békésy (1899-1972) verwendete ein Mikroskop, um die im Innenohr von Leichen befindliche Basilarmembran zu untersuchen. Er fand heraus, dass die Bewegung der Basilarmembran der einer Wanderwelle ähnelt; Die Form variiert je nach Frequenz der Tonhöhe. Bei niederfrequenten Geräuschen bewegt sich die Spitze (Spitze) der Membran am meisten, während sich bei hochfrequenten Geräuschen die Basis der Membran am meisten bewegt.
Störungen
Eine Störung oder Infektion des Labyrinths kann zu einem Krankheitssyndrom führen, das als Labyrinthitis bezeichnet wird . Die Symptome einer Labyrinthitis sind vorübergehende Übelkeit, Orientierungslosigkeit, Schwindel und Schwindel. Labyrinthitis kann durch Virusinfektionen, bakterielle Infektionen oder körperliche Blockaden des Innenohrs verursacht werden.
Eine andere Erkrankung ist als Autoimmunerkrankung des Innenohrs (AIED) bekannt geworden. Es ist gekennzeichnet durch einen idiopathischen, schnell fortschreitenden bilateralen sensorineuralen Hörverlust. Es ist eine ziemlich seltene Erkrankung, und gleichzeitig hat das Fehlen geeigneter diagnostischer Tests dazu geführt, dass die genaue Inzidenz nicht bestimmt werden kann.
Andere Tiere
Vögel haben ein ähnliches Hörsystem wie Säugetiere, einschließlich einer Cochlea. Reptilien, Amphibien und Fische haben keine Cochleas, sondern hören mit einfacheren Hörorganen oder Vestibularorganen, die im Allgemeinen niederfrequente Geräusche als die Cochlea erkennen. Die Cochlea von Vögeln ähnelt der von Krokodilen und besteht aus einem kurzen, leicht gebogenen Knochenrohr, in dem die Basilarmembran mit ihren sensorischen Strukturen liegt.
Cochlea-System
Bei Reptilien wird der Schall vom Stapes (Steigbügel) des Mittelohrs auf das Innenohr übertragen. Dies wird gegen das ovale Fenster gedrückt , eine membranbedeckte Öffnung auf der Oberfläche des Vestibüls. Von hier aus werden Schallwellen durch einen kurzen perilymphatischen Kanal zu einer zweiten Öffnung geleitet, dem runden Fenster , das den Druck ausgleicht und es der inkompressiblen Flüssigkeit ermöglicht, sich frei zu bewegen. Parallel zum Ductus perilymphaticus verläuft ein separater Ductus mit blindem Ende, die Lagena , die mit Endolymphe gefüllt ist . Die Lagena ist durch eine Basilarmembran vom Ductus perilymphaticus getrennt und enthält die sensorischen Haarzellen, die die Schwingungen in der Flüssigkeit schließlich in Nervensignale umwandeln. Es ist an einem Ende am Sacculus befestigt.
Bei den meisten Reptilien sind der perilymphatische Gang und die Lagena relativ kurz, und die Sinneszellen sind auf eine kleine Papille beschränkt, die zwischen ihnen liegt. Bei Vögeln , Säugetieren und Krokodilen werden diese Strukturen jedoch viel größer und etwas komplizierter. Bei Vögeln, Krokodilen und Monotremen sind die Kanäle einfach verlängert und bilden zusammen ein längliches, mehr oder weniger gerades Rohr. Der endolymphatische Gang ist in einer einfachen Schleife um die Lagena gewickelt, wobei die Basilarmembran an einer Seite liegt. Die erste Hälfte des Ganges wird jetzt als Scala Vestibuli bezeichnet , während die zweite Hälfte, die die Basilarmembran enthält, als Scala Tympani bezeichnet wird . Infolge dieser Längenzunahme werden sowohl die Basilarmembran als auch die Papille verlängert, wobei sich letztere zum Corti-Organ entwickelt , während die Lagena jetzt als Cochlea-Gang bezeichnet wird . Alle diese Strukturen bilden zusammen die Cochlea.
Bei Säugetieren (außer Monotremen) ist die Cochlea noch weiter verlängert und wird zu einer gewundenen Struktur, um ihre Länge im Kopf aufzunehmen. Das Corti-Organ hat auch bei Säugetieren eine komplexere Struktur als bei anderen Amnioten .
Die Anordnung des Innenohrs bei lebenden Amphibien ähnelt in vielerlei Hinsicht der von Reptilien. Oft fehlt ihnen jedoch eine Basilarpapille, die stattdessen einen völlig getrennten Satz von Sinneszellen am oberen Rand des Sacculus aufweist, der als Papilla amphibiorum bezeichnet wird und die gleiche Funktion zu haben scheint.
Obwohl viele Fische hörfähig sind, ist die Lagena bestenfalls ein kurzes Divertikel des Sacculus und scheint keine Rolle bei der Klangempfindung zu spielen. Stattdessen können verschiedene Haarzellencluster im Innenohr verantwortlich sein. Beispielsweise enthalten knöcherne Fische im Utrikel einen sensorischen Cluster, der als Macula Neglecta bezeichnet wird und diese Funktion haben kann. Obwohl Fische weder ein Außen- noch ein Mittelohr haben, kann der Schall dennoch über die Schädelknochen oder über die Schwimmblase auf das Innenohr übertragen werden , von denen Teile häufig in der Nähe des Körpers liegen.
Vestibularsystem
Im Vergleich zum Cochlea- System variiert das Vestibularsystem zwischen den verschiedenen Gruppen von Kieferwirbeltieren relativ wenig . Der zentrale Teil des Systems besteht aus zwei Kammern, dem Sacculus und dem Utrikel, von denen jede ein oder zwei kleine Cluster sensorischer Haarzellen enthält. Alle Wirbeltiere mit Kiefer besitzen außerdem drei halbkreisförmige Kanäle, die aus dem Utrikel hervorgehen und an deren einem Ende jeweils eine Ampulle mit Sinneszellen vorhanden ist.
Ein endolymphatischer Gang verläuft vom Sacculus durch den Kopf und endet nahe am Gehirn. Bei knorpeligen Fischen öffnet sich dieser Kanal tatsächlich zur Oberseite des Kopfes, und bei einigen Teleost- Fischen ist er einfach blind. Bei allen anderen Arten endet es jedoch in einem endolymphatischen Sack . Bei vielen Reptilien, Fischen und Amphibien kann dieser Sack eine beträchtliche Größe erreichen. Bei Amphibien können die Säcke von beiden Seiten zu einer einzigen Struktur verschmelzen, die sich häufig parallel zum Wirbelkanal über die gesamte Körperlänge erstreckt .
Die primitiven Neunaugen und Inger haben jedoch ein einfacheres System. Das Innenohr dieser Art besteht aus einer einzigen Vestibularkammer, obwohl dies bei Neunaugen mit einer Reihe von von Zilien ausgekleideten Säcken verbunden ist . Neunaugen haben nur zwei halbkreisförmige Kanäle, wobei der horizontale Kanal fehlt, während Hagfish nur einen einzigen vertikalen Kanal hat.
Gleichgewicht
Das Innenohr ist in erster Linie für das Gleichgewicht, das Gleichgewicht und die Orientierung im dreidimensionalen Raum verantwortlich. Das Innenohr kann sowohl das statische als auch das dynamische Gleichgewicht erfassen. Drei halbkreisförmige Kanäle und zwei Kammern, die den Sacculus und den Utrikel enthalten , ermöglichen es dem Körper, Abweichungen vom Gleichgewicht festzustellen. Der Macula sacculi erkennt die vertikale Beschleunigung, während der Macula utriculi für die horizontale Beschleunigung verantwortlich ist. Diese mikroskopischen Strukturen besitzen Stereozilien und ein Kinozilium, die sich innerhalb der gelatinösen otolithischen Membran befinden. Die Membran wird weiter mit Otolithen gewichtet. Durch die Bewegung der Stereozilien und des Kinoziliums können die Haarzellen der Saccula und des Utrikels Bewegungen erkennen. Die halbkreisförmigen Kanäle sind für die Erfassung der Drehbewegung verantwortlich.
Zusätzliche Bilder

Ohrlabyrinth

Innenohr

Schläfenbein
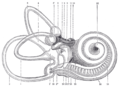
Rechtes menschliches membranöses Labyrinth, entfernt von seiner knöchernen Hülle und von antero-lateral betrachtet





Siehe auch
Verweise
- Ruckenstein, MJ (2004). "Autoimmun-Innenohr-Krankheit". Current Opinion in Otolaryngology & Head and Neck Surgery, 12 (5), S. 426–430.
Saladin, "Anatomy and Physiology" 6e, Print American Speech-Language-Hearing Association, Mittelohr, https://web.archive.org/web/20131208002201/http://www.asha.org/public/hearing/ Mittelohr/
Externe Links
Anatomiefoto: 30: 05-0101 im SUNY Downstate Medical Center