
Epithelioidzelle - Epithelioid cell

Gemäß einer gemeinsamen Sicht Epitheloidzellen (auch genannt epithelioid Histiozyten ) sind Derivate von aktivierten Makrophagen ähnlich Epithelzellen .
Struktur und Funktion

Strukturell sind epitheloide Zellen (bei lichtmikroskopischer Untersuchung nach Färbung mit Hämatoxylin und Eosin) länglich, mit feinkörnigem, blassem eosinophilem (rosa) Zytoplasma und zentralen, eiförmigen Kernen (oval oder länglich), die weniger dicht sind als bei ein Lymphozyt . Sie haben eine undeutliche Form und scheinen oft ineinander zu verschmelzen und bilden Aggregate, die als Riesenzellen bekannt sind. Bei der Transmissionselektronenmikroskopie an epitheloiden Zellen im Bereich des Golgi-Lamellenkomplexes werden nicht nur zonierte, sondern auch glatte Vesikel mit dichtem Zentrum, sowie sehr viele (über 100) große Granula mit Durchmessern bis 340 nm und mit feinkörnigen Matrix heller als in Makrophagen-Granula, manchmal mit perigranulärem Halo. „Das auffälligste Merkmal dieser Zellen ist das riesige Golgi-Gebiet; bis zu 6 einzelne Stapel von Golgi-Zisternen können vorhanden sein sowie einige wenige borstenüberzogene und zahlreiche glatte Bläschen“. Epithelioide Zellen haben eng ineinandergreifende Zellmembranen in reißverschlussartigen Anordnungen, die benachbarte Zellen verbinden. Diese Zellen sind von zentraler Bedeutung für die Bildung von Granulomen, die mit vielen schweren Krankheiten in Verbindung gebracht werden. Bei Granulomen erfüllen Epitheloidzellen die Funktionen der Abgrenzung.
Besonderheiten des Zytoskeletts

Es wird gezeigt, dass sich das durch Filamente gebildete epitheloide Zellzytoskelett signifikant vom Makrophagenzytoskelett unterscheidet. Eine starke Zunahme der Filamentzahl tritt in diesen Zellen auf, wobei Filamente (90 bis 100 A) das Zytozentrum als markantes ringförmiges Bündel umgeben, das sich oft in das Zytoplasma verzweigt. Aufgrund eines solchen Zytoskeletts weisen angrenzende Epitheloidzellen eine ausgeklügelte zytoplasmatische Verflechtung auf. Durch die Verwendung der Quick=Freeze- und Freeze-Substitution-Methoden (promptes Freeze, penetrierendes Ätzen und Freeze-Substitution) wurde gezeigt, dass die Organisationen dreidimensionale Metastruktur des Zytoskeletts der Epitheloidzellen, die im Fokus der granulomatösen Entzündung gebildet werden , besser kompatibel zu Zytoskelett, das für typische Epithelzellen charakteristisch ist, als für das Zytoskelett von aktiven und beweglichen Makrophagen. Es wird gezeigt, dass die dichten Netze intermediärer Filamente, die mit Kernen, Mitochondrien und anderen Organellen verbunden sind, überall im Zytoplasma von Epitheloidzellen überwacht werden. Einige Faszikel aus aktinischen Filamenten wurden in Filopodien unterhalb der Membranen der Zellen angeordnet. Eine exakte interdigitale Verdreifachung von Zellmembranen zwischen aneinandergrenzenden Epitheloidzellen wurde klar demonstriert. Interdigitale Filopodien wurden identifiziert. Das charakteristische Anzeichen von Epitheloidzellen ist ihre Aggregation mit Bildung enger interdigitaler Verdreifachungen als Befestigungs "Blitz", der anscheinend die Bedeutung für die Bildung einer dichten Abgrenzungszone des Körpers vom Erreger während der Bildung von Epitheloidzellen haben kann Granulome.
Immunologischer Phänotyp und immunologische Marker
Bei der Verwendung von Antikörpern gegen die RFD9-, RFD7- und HLA-DR-Antigene wurde festgestellt, dass alle Epitheloidzellen einen immunologischen Phänotyp RFD9+/RFD7-/HLA-DR+ aufweisen. Es wurde eine Reihe von monoklonalen Antikörpern IHY-1, IHY-2, IHY-3 erhalten, die verwendet werden können, um Epithelzellen, die bei ätiologisch unterschiedlichen Formen der granulomatösen Entzündung gebildet wurden, genau zu identifizieren. Der IHY-1-Antikörper reagiert mit Epitheloidzellen in Sarkoid- Granulomen sowie mit Epitheloidzellen verschiedener granulomatöser Erkrankungen, einschließlich Tuberkulose. IHY-2 und IHY-3 l Antikörper, monoklonaler Antikörper, reagieren mit Epitheloidzellen bei Sarkoidose, aber nicht bei Tuberkulose .
Klinische Bedeutung
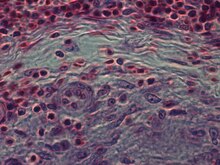
Epithelioide Zellen sind ein wesentliches Merkmal von epitheloiden Zellgranulomen . Epithelioidzell-Granulome können als spezifisch und strukturell organisierte Ansammlungen von Epitheloidzellen, Makrophagen, Lymphozyten und dendritischen Zellen definiert werden. Fremdkörpergranulome können als organisierte Ansammlung von Makrophagen angesehen werden, einschließlich bloßer Ansammlungen von Riesenzellen, die inerte Substanzen wie Nahtmaterial umgeben – die sogenannten "nicht-immunen Granulome". Die Bildung von Granulomen wird mit Krankheitserregern in Verbindung gebracht, die gelernt haben, dem Immunsystem des Wirts durch verschiedene Maßnahmen zu entgehen, wie z. B. der Phagozytose zu widerstehen und innerhalb der Makrophagen abzutöten. Die Unverdaulichkeit von Stoffen durch Makrophagen ist ein häufiges Merkmal einer granulomatösen Entzündung. Granulome versuchen, diese Organismen abzuschirmen und ihr weiteres Wachstum und ihre Ausbreitung zu verhindern. Historisch weit verbreitete und destruktive Erkrankungen wie Tuberkulose, Lepra und Syphilis sind granulomatöse Erkrankungen. Die Bildung von Granulomen ist auch das Merkmal vieler modernerer Erkrankungen, wie Pilzinfektionen, Sarkoidose und Morbus Crohn .
Geschichte der wissenschaftlichen Forschung
Die erste Erwähnung von Epitheloidzellen als spezifischer Zellform erfolgte im 19. Jahrhundert in Werken von Koch R. und Cornil J., die glaubten, dass die Leukozyten die Urheber der Epitheloidzellen der Tuberkulose seien. Yersin A (1888) und Borrel A (1893) zeigten in Versuchen an Kaninchen, dass Epitheloidzellen aus mononukleären Blutleukozyten gebildet werden. Die Hauptmuster der Bildung von Epitheloidzellen wurden erstmals in der ersten Hälfte des 20. Jahrhunderts von Lewis M. (1925) beschrieben. Dieser Forscher zeigte, dass Blutmonozyten in Zellkulturen von Mischblut-Leukozyten von Vögeln (von ausgewachsenen Hühnern sowie von Embryonen unterschiedlichen Alters), Mäusen und Menschen, wenn sie in vitro kultiviert werden, in typische Makrophagen und epitheloide Zellen umgewandelt werden, gefolgt durch die Bildung von riesigen mehrkernigen Zellen. Die Bildung von Zellen vom Epitheloidtyp wurde von Lewis M. am 2.-3. Tag der Kultivierung von Leukozyten festgestellt. Später in einer Studie eines ähnlichen Plans zeigten Jerry S und Weiss L (1966), als sie Kulturen von Mischblut-Leukozyten von Hühnern (getrennt aus Herzblut von Rhode Island Red) und Elektronenmikroskopie verwendeten, dass die Transformation von Hühnermonozyten in Epitheloidzellen beginnen in Kultur nach 3–4 Tagen und enden nach 5–6 Tagen. Da alle früheren Forscher darauf hingewiesen haben, dass Epitheloidzellen aus Monozyten gebildet werden und Monozyten und Makrophagen zu einem einzigen mononuklearen Phagozytensystem kombiniert wurden , haben Van Furth et al. (1972) unter Bezugnahme auf die Arbeiten von Sutton J. und Weiss L. (1966) schrieben epitheloide Zellen formal dem mononuklearen Phagozytensystem zu. Sie gaben jedoch nicht genau an, aus welchen Zellen des mononukleären Phagozytensystems epitheloide Zellen stammen. Gleichzeitig zogen sie den sehr vorsichtigen Schluss: «Die in diesen Läsionen vorkommenden Epitheloidzellen stammen ebenfalls aus Monozyten oder Makrophagen». Adams D (1976), der glaubte, dass Epitheloidzellen die letzte Stufe der Zelldifferenzierung der Zellen des mononuklearen Phagozytensystems sind, formulierte das Konzept der Zytomorphogeese von Epitheloidzellen, nach dem Epitheloidzellen als Derivat aktivierter Makrophagen angesehen werden (was wird derzeit noch von den meisten Forschern gehalten). Dies basiert auf der Annahme, dass "die Stimulierung von Makrophagen weiter zu unreifen Epitheloidzellen und schließlich zu reifen reifen". Rhee et al. (1979) in Experimenten an Ratten mit der Methode der Elektronenmikroskopie gezeigt, dass eines der wichtigsten zytomorphologischen Merkmale von Epitheloidzellen, die diese Zellen von Makrophagen unterscheiden, das Vorhandensein charakteristischer spezifischer Granula in ihnen ist, die sie Epitheloidzellgranula nannten. Basierend auf ihren eigenen Daten unterstützten sie das Konzept der Zytomorphogenese von Epitheloidzellen, nach dem Epitheloidzellen als Derivat eines aktivierten Makrophagen angesehen werden. Später schlugen Turk JL und Narayanan RB (1982) vor, in der Studie zwei Arten von Epitheloidzellen zu unterscheiden : "vesikuläre" und "sekretorische" Epitheloidzellen. Es wird vermutet, dass sich „vesikuläre Epitheloidzellen aus „sekretorischen“ Epitheloidzellen durch einen Degenerationsprozess entwickeln könnten. Auf der Suche nach immunologischen Mechanismen, die die Bildung von Epithelzellen beeinflussen, erhielten Cipriano et al. (2003) Daten, die einen möglichen Einfluss von IL- 4 über die Bildung eines Phänotyps in Makrophagen, der dem Phänotyp von Epitheloidzellen ähnlich ist.Allerdings passen nicht alle Ergebnisse der Forschungen zu den Gesetzen und Mechanismen der Zytomorphogenese von Epitheloidzellen in das Konzept des Ursprungs von Epitheloidzellen aus Makrophagen Deimann J und Fahimi H (1980) zeigten, dass epitheloide Zellen in Granulomen, die in der Rattenleber durch Injektion von Glucan beta-1,30-Polyglucose induziert wurden, nicht aus Kupffer-Zellen - reifen differenzierten Makrophagen, sondern aus Blut gebildet werden Monozyten. De Vos et al. (1990) erhalten die Daten , die sie erlaubt , dass Brennpunkte in granulomatöse Entzündung vorschlagen, in granulomatöse Lymphadenitis , Epitheloidzellen nicht aus differenzierten Makrophagen gebildet werden, sondern aus sogenannten plasmazytoiden Monozyten (haben Ähnlichkeiten mit Plasmozyten). Dies wird weiter durch die ultrastrukturellen Ähnlichkeiten zwischen plasmazytoiden Monozyten und epitheloiden Zellen unterstützt. Die vorliegende ultrastrukturelle und immunelektronenmikroskopische Untersuchung von Epitheloidzellgranulomen lieferte weitere Argumente für diese Hypothese. Arkhipov S (1997, 2012) zeigte anhand von Kulturen von Peritonealzellen, Blutleukozyten und Knochenmarkszellen von Mäusen, dass Makrophagen und Epitheloidzellen aus verschiedenen Monozytentypen gebildet werden. Es wurde gezeigt, dass Epitheloidzellen nur aus Monozyten vom plasmozytoiden Typ gebildet werden, die als Präepitheloidzellen bezeichnet werden, wobei das Stadium der Differenzierung zu Makrophagen umgangen wird . Es hat sich gezeigt, dass bei chronischen Entzündungen die Zahl der präepitheloiden Monozyten, die an der epitheloiden Zelldifferenzierung beteiligt sind, im Entzündungsherd, im Blut und im Knochenmark, zunimmt. unter Verwendung von Maus- Inzuchtlinien , im Gegensatz zur Anfälligkeit für Mycobacterium tuberculosis , wurde gezeigt, dass die Anzahl der präepitheloiden Monozyten, die bei chronischen Entzündungen gebildet werden, genetisch bestimmt ist. Die erhaltenen Ergebnisse zeigten, dass die Morphogenese von epitheloidzelligen Granulomen durch die unterschiedlichen genetisch bedingten Ausgangsniveaus eines Pools von präepitheloiden Zellen des monozytoiden Typs, deren Fluss in das Zentrum einer granulomatösen Entzündung, die Intensität der Differenzierung in epitheloide Zellen, Umgehen des Stadiums der Differenzierung in Makrophagen und ihre endomitotische Aktivität.
Siehe auch
Verweise
Externe Links
- Epithelioid+Zellen in der US National Library of Medicine Medical Subject Headings (MeSH)
Dieser zellbiologische Artikel ist ein Stummel . Sie können Wikipedia helfen, indem Sie es erweitern . |